



អង្គបដិប្រាណដែលត្រូវបានគេហៅថា immunoglobulins (Ig) គឺជា glycoproteins ដែលភ្ជាប់ជាពិសេសទៅនឹងអង់ទីហ្សែន។
ការរៀបចំអង្គបដិប្រាណធម្មតាត្រូវបានផលិតដោយការចាក់ថ្នាំបង្ការសត្វ និងការប្រមូលថ្នាំប្រឆាំងនឹងសេរ៉ូម។ដូច្នេះ antiserum ជាធម្មតាមានអង្គបដិប្រាណប្រឆាំងនឹង antigens ដែលមិនទាក់ទងនិងសមាសធាតុប្រូតេអ៊ីនផ្សេងទៀតនៅក្នុងសេរ៉ូម។ម៉ូលេគុលអង់ទីហ្សែនទូទៅភាគច្រើនមានអេពីតូបផ្សេងៗគ្នា ដូច្នេះអង្គបដិប្រាណធម្មតាក៏ជាល្បាយនៃអង្គបដិប្រាណប្រឆាំងនឹងអេពីតូបផ្សេងៗគ្នាជាច្រើនផងដែរ។សូម្បីតែអង្គបដិប្រាណសេរ៉ូមធម្មតាដែលដឹកនាំប្រឆាំងនឹងអេពីតូបដូចគ្នា នៅតែត្រូវបានផ្សំឡើងដោយអង្គបដិប្រាណផ្សេងៗគ្នាដែលផលិតដោយក្លូនកោសិកា B ផ្សេងៗគ្នា។ដូច្នេះ អង្គបដិប្រាណសេរ៉ូមធម្មតា ត្រូវបានគេហៅផងដែរថា អង្គបដិបក្ខ polyclonal ឬ អង្គបដិប្រាណ polyclonal សម្រាប់រយៈពេលខ្លី។
អង្គបដិបក្ខ Monoclonal (អង្គបដិបក្ខ monoclonal) គឺជាអង្គបដិប្រាណដែលមានឯកសណ្ឋានខ្ពស់ដែលផលិតដោយក្លូនកោសិកា B តែមួយ ហើយដឹកនាំតែប្រឆាំងនឹងអេពីតូបជាក់លាក់មួយ។ជាធម្មតាវាត្រូវបានរៀបចំដោយបច្ចេកវិទ្យា hybridoma - បច្ចេកវិទ្យាអង្គបដិប្រាណ hybridoma គឺផ្អែកលើបច្ចេកវិទ្យានៃការបញ្ចូលគ្នានៃកោសិកា ដោយរួមបញ្ចូលគ្នានូវកោសិកា B ជាមួយនឹងសមត្ថភាពក្នុងការបញ្ចេញអង្គបដិប្រាណជាក់លាក់ និងកោសិកា myeloma ជាមួយនឹងសមត្ថភាពលូតលាស់គ្មានកំណត់ទៅជាកូនកាត់កោសិកា B ។កោសិកាកូនកាត់នេះមានលក្ខណៈនៃកោសិកាមេ។វាអាចរីកសាយភាយដោយគ្មានកំណត់ និងជាអមតៈនៅក្នុង vitro ដូចជាកោសិកា myeloma ហើយវាអាចសំយោគ និងសម្ងាត់អង្គបដិប្រាណជាក់លាក់ដូចជា lymphocytes splenic ។តាមរយៈការក្លូន ខ្សែ monoclonal ដែលកើតចេញពីកោសិកា hybridoma តែមួយ នោះគឺ ខ្សែកោសិកា hybridoma អាចទទួលបាន។អង្គបដិប្រាណដែលវាផលិតគឺជាអង្គបដិប្រាណដូចគ្នាយ៉ាងខ្ពស់ប្រឆាំងនឹងកត្តាកំណត់អង់ទីហ្សែនដូចគ្នា ពោលគឺអង្គបដិប្រាណ monoclonal ។
អង្គបដិបក្ខមានជាម៉ូណូម័ររាងអក្សរ Y មួយ ឬច្រើន (ឧទាហរណ៍ អង្គបដិបក្ខ monoclonal ឬអង្គបដិបក្ខ polyclonal) ។ម៉ូណូម័ររាងអក្សរ Y នីមួយៗមានខ្សែសង្វាក់ polypeptide ចំនួន 4 រួមទាំងខ្សែសង្វាក់ធ្ងន់ដូចគ្នាចំនួនពីរ និងខ្សែសង្វាក់ពន្លឺដូចគ្នាបេះបិទ។ខ្សែសង្វាក់ស្រាល និងខ្សែសង្វាក់ធ្ងន់ត្រូវបានដាក់ឈ្មោះតាមទម្ងន់ម៉ូលេគុលរបស់វា។ផ្នែកខាងលើនៃរចនាសម្ព័ន្ធរាងអក្សរ Y គឺជាតំបន់អថេរ ដែលជាកន្លែងភ្ជាប់អង់ទីហ្សែន។(ដកស្រង់ចេញពី ទស្សនៈ ឌីតៃ ជីវ-ម៉ូណូក្លូន អង់ទីគ័រ)
រចនាសម្ព័ន្ធអង្គបដិប្រាណ
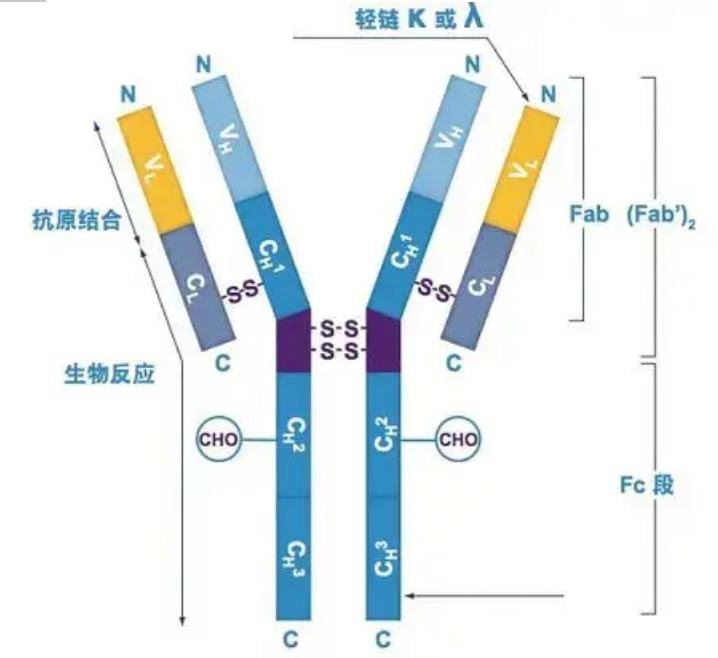 ខ្សែសង្វាក់ធ្ងន់
ខ្សែសង្វាក់ធ្ងន់
មានខ្សែសង្វាក់ធុនធ្ងន់ Ig ថនិកសត្វចំនួនប្រាំប្រភេទ ដែលដាក់ឈ្មោះជាអក្សរក្រិច α, δ, ε, γ និង μ។អង្គបដិប្រាណដែលត្រូវគ្នាត្រូវបានគេហៅថា IgA, IgD, IgE, IgG និង IgM ។ខ្សែសង្វាក់ធ្ងន់ខុស ៗ គ្នាមានទំហំនិងសមាសភាព។α និង γ មានអាស៊ីតអាមីណូប្រហែល 450 ខណៈពេលដែល μ និង ε មានអាស៊ីតអាមីណូប្រហែល 550 ។
ខ្សែសង្វាក់ធ្ងន់នីមួយៗមានតំបន់ពីរ៖ តំបន់ថេរ និងតំបន់អថេរ។អង្គបដិប្រាណទាំងអស់នៃប្រភេទដូចគ្នាមានតំបន់ថេរដូចគ្នា ប៉ុន្តែមានភាពខុសគ្នារវាងអង្គបដិប្រាណនៃប្រភេទផ្សេងៗគ្នា។តំបន់ថេរនៃខ្សែសង្វាក់ធ្ងន់ γ, α, និង δ ត្រូវបានផ្សំឡើងដោយដែន Ig បីស្របជាមួយនឹងតំបន់ hinge ដើម្បីបង្កើនភាពបត់បែនរបស់វា;តំបន់ថេរនៃខ្សែសង្វាក់ធ្ងន់ μ និង ε ត្រូវបានផ្សំឡើងដោយដែន 4 Ig ។តំបន់អថេរនៃខ្សែសង្វាក់ធ្ងន់នៃអង្គបដិប្រាណដែលផលិតដោយកោសិកា B ផ្សេងគ្នាគឺខុសគ្នា ប៉ុន្តែតំបន់អថេរនៃអង្គបដិប្រាណដែលផលិតដោយកោសិកា B ឬក្លូនកោសិកាដូចគ្នាគឺដូចគ្នា ហើយតំបន់អថេរនៃខ្សែសង្វាក់ធ្ងន់នីមួយៗមានប្រវែងប្រហែល 110 អាមីណូអាស៊ីត។និងបង្កើតដែន Ig តែមួយ។
ខ្សែសង្វាក់ពន្លឺ
មានខ្សែសង្វាក់ពន្លឺពីរប្រភេទនៅក្នុងថនិកសត្វ: ប្រភេទ lambda និងប្រភេទ kappa ។ខ្សែសង្វាក់ពន្លឺនីមួយៗមានដែនតភ្ជាប់ពីរ៖ តំបន់ថេរ និងតំបន់អថេរ។ប្រវែងនៃខ្សែសង្វាក់ពន្លឺគឺប្រហែល 211 ~ 217 អាស៊ីតអាមីណូ។ខ្សែសង្វាក់ពន្លឺពីរដែលមាននៅក្នុងអង្គបដិប្រាណនីមួយៗគឺតែងតែដូចគ្នា។សម្រាប់ថនិកសត្វ ខ្សែសង្វាក់ពន្លឺនៅក្នុងអង្គបដិប្រាណនីមួយៗមានតែមួយប្រភេទប៉ុណ្ណោះ៖ កាប៉ា ឬឡាំដា។នៅក្នុងសត្វឆ្អឹងកងទាបមួយចំនួនដូចជា ត្រីឆ្អឹងខ្ចី (ត្រីឆ្អឹងខ្ចី) និងត្រីឆ្អឹង ប្រភេទខ្សែសង្វាក់ពន្លឺផ្សេងទៀតដូចជាប្រភេទ iota (iota) ក៏ត្រូវបានរកឃើញផងដែរ។
ផ្នែក Fab និង Fc
ផ្នែក Fc អាចត្រូវបានផ្សំដោយផ្ទាល់ជាមួយអង់ស៊ីម ឬថ្នាំជ្រលក់ fluorescent ដើម្បីដាក់ស្លាកអង្គបដិប្រាណ។វាគឺជាផ្នែកដែលអង្គបដិប្រាណចងជាប់នៅលើចានកំឡុងពេលដំណើរការ ELISA ហើយវាក៏ជាផ្នែកដែលអង្គបដិប្រាណទីពីរត្រូវបានទទួលស្គាល់ និងចងនៅក្នុង immunoprecipitation, immunoblotting និង immunohistochemistry ។អង្គបដិប្រាណអាចត្រូវបានបំប្លែងទៅជាផ្នែក F(ab) ពីរ និងផ្នែក Fc មួយដោយអង់ស៊ីម proteolytic ដូចជា papain ឬពួកវាអាចត្រូវបានបំបែកចេញពីតំបន់ hinge ដោយ pepsin និង hydrolyzed ទៅជា F(ab)2 segment និង Fc មួយផ្នែក។បំណែកអង្គបដិប្រាណ IgG ជួនកាលមានប្រយោជន៍ខ្លាំងណាស់។ដោយសារកង្វះផ្នែក Fc ផ្នែក F(ab) នឹងមិនជ្រាបចូលជាមួយអង់ទីហ្សែនទេ ហើយក៏មិនត្រូវបានចាប់យកដោយកោសិកាភាពស៊ាំនៅក្នុងការសិក្សា vivo ដែរ។ដោយសារតែបំណែកម៉ូលេគុលតូច និងកង្វះមុខងារតំណភ្ជាប់ឆ្លងកាត់ (ដោយសារតែខ្វះផ្នែក Fc) ផ្នែក Fab ជាធម្មតាត្រូវបានប្រើសម្រាប់ការដាក់ស្លាកវិទ្យុក្នុងការសិក្សាមុខងារ ហើយផ្នែក Fc ត្រូវបានប្រើជាចម្បងជាភ្នាក់ងារទប់ស្កាត់ក្នុងការស្នាមប្រឡាក់គីមី។
តំបន់អថេរ និងថេរ
តំបន់អថេរ (តំបន់ V) មានទីតាំងនៅ 1/5 ឬ 1/4 (មានសំណល់អាស៊ីតអាមីណូប្រហែល 118) នៃខ្សែសង្វាក់ H នៅជិត N-terminus និង 1/2 (មានសំណល់អាស៊ីតអាមីណូប្រហែល 108-111) នៅជិត N-terminus នៃខ្សែសង្វាក់ L ។តំបន់ V នីមួយៗមានរង្វង់ peptide ដែលបង្កើតឡើងដោយចំណង disulfide ខាងក្នុងខ្សែសង្វាក់ ហើយចិញ្ចៀន peptide នីមួយៗមានសំណល់អាស៊ីតអាមីណូប្រហែល 67 ទៅ 75 ។សមាសភាព និងការរៀបចំអាស៊ីតអាមីណូនៅក្នុងតំបន់ V កំណត់ភាពជាក់លាក់នៃការភ្ជាប់អង់ទីហ្សែននៃអង្គបដិប្រាណ។ដោយសារប្រភេទ និងលំដាប់នៃអាស៊ីតអាមីណូដែលផ្លាស់ប្តូរជានិច្ចនៅក្នុងតំបន់ V អង្គបដិប្រាណជាច្រើនប្រភេទដែលមានភាពជាក់លាក់អង់ទីហ្សែនចងផ្សេងៗគ្នាអាចត្រូវបានបង្កើតឡើង។តំបន់ V នៃខ្សែសង្វាក់ L និង H ត្រូវបានគេហៅថា VL និង VH រៀងគ្នា។នៅក្នុង VL និង VH សមាសធាតុអាស៊ីតអាមីណូ និងលំដាប់នៃតំបន់មួយចំនួនមានកម្រិតនៃការប្រែប្រួលខ្ពស់ជាង។តំបន់ទាំងនេះត្រូវបានគេហៅថាតំបន់អថេរខ្ពស់ (HVR) ។សមាសធាតុអាស៊ីតអាមីណូ និងការរៀបចំផ្នែកដែលមិនមែនជា HVR នៅក្នុងតំបន់ V គឺមានលក្ខណៈអភិរក្ស ដែលត្រូវបានគេហៅថាតំបន់ក្របខ័ណ្ឌ។មានតំបន់អថេរបីនៅក្នុង VL ដែលជាធម្មតាមានទីតាំងនៅសំណល់អាស៊ីតអាមីណូ 24 ទៅ 34 និង 89 ទៅ 97 រៀងគ្នា។HVRs បីនៃ VL និង VH ត្រូវបានគេហៅថា HVR1, HVR2 និង HVR3 រៀងគ្នា។ការស្រាវជ្រាវ និងការវិភាគនៃការបំភាយគ្រីស្តាល់កាំរស្មីអ៊ិច បានបង្ហាញថា តំបន់អថេរខ្ពស់គឺពិតជាកន្លែងដែលអង់ទីករអង់ទីគ័រចង ដូច្នេះគេហៅថាតំបន់កំណត់ការបំពេញបន្ថែម (CDR)។HVR1, HVR2 និង HVR3 នៃ VL និង VH អាចត្រូវបានគេហៅថា CDR1, CDR2 និង CDR3 រៀងគ្នា។ជាទូទៅ CDR3 មានកម្រិតខ្ពស់នៃភាពប្រែប្រួលខ្ពស់។តំបន់អថេរខ្ពស់ក៏ជាទីតាំងសំខាន់ដែលកត្តាកំណត់អត្តសញ្ញាណនៃម៉ូលេគុល Ig មាន។ក្នុងករណីភាគច្រើន ខ្សែសង្វាក់ H មានតួនាទីសំខាន់ជាងក្នុងការភ្ជាប់ទៅនឹងអង់ទីហ្សែន។
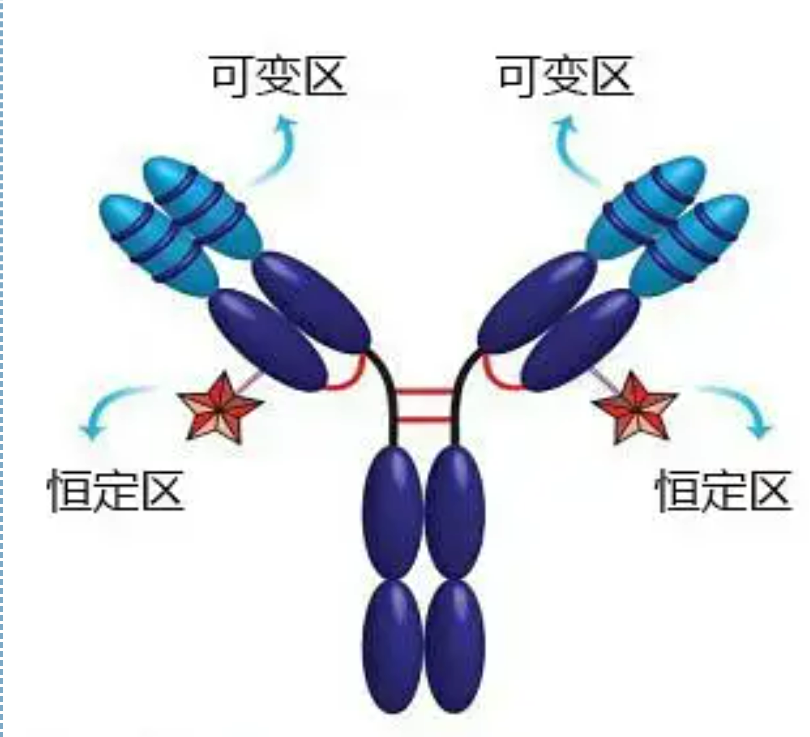 តំបន់ថេរ (C តំបន់)មានទីតាំងនៅ 3/4 ឬ 4/5 (ប្រហែលពីអាស៊ីតអាមីណូ 119 ទៅស្ថានីយ C) នៃខ្សែសង្វាក់ H នៅជិតស្ថានីយ C និង 1/2 (មានសំណល់អាស៊ីតអាមីណូប្រហែល 105) នៅជិតស្ថានីយ C នៃខ្សែសង្វាក់ L ។តំបន់មុខងារនីមួយៗនៃខ្សែសង្វាក់ H មានសំណល់អាស៊ីតអាមីណូប្រហែល 110 ហើយមានចិញ្ចៀន peptide ដែលផ្សំឡើងដោយសំណល់អាស៊ីតអាមីណូ 50-60 ដែលតភ្ជាប់ដោយចំណង disulfide ។សមាសធាតុអាស៊ីតអាមីណូ និងការរៀបចំនៃតំបន់នេះគឺថេរក្នុងខ្សែសង្វាក់ Ig isotype L សត្វដូចគ្នា និងខ្សែសង្វាក់ H ប្រភេទដូចគ្នា។ដូចគ្នាដែរ វាអាចភ្ជាប់ជាពិសេសទៅនឹងអង់ទីហ្សែនដែលត្រូវគ្នា ប៉ុន្តែរចនាសម្ព័ន្ធនៃតំបន់ C របស់វាគឺដូចគ្នា ពោលគឺវាមានអង់ទីហ្សែនដូចគ្នា។អង់ទីករបន្ទាប់បន្សំ IgG ប្រឆាំងនឹងមនុស្ស (ឬប្រឆាំងនឹងអង្គបដិប្រាណ) អាចត្រូវបានផ្សំជាមួយនឹងការរួមបញ្ចូលគ្នានៃអង្គបដិប្រាណ (IgG) ពីរប្រឆាំងនឹង exotoxins ផ្សេងគ្នាកើតឡើង។នេះគឺជាមូលដ្ឋានដ៏សំខាន់សម្រាប់ការរៀបចំអង្គបដិប្រាណបន្ទាប់បន្សំ និងអនុវត្ត fluorescein អ៊ីសូតូប អង់ស៊ីម និងអង្គបដិប្រាណដែលមានស្លាកផ្សេងទៀត។
តំបន់ថេរ (C តំបន់)មានទីតាំងនៅ 3/4 ឬ 4/5 (ប្រហែលពីអាស៊ីតអាមីណូ 119 ទៅស្ថានីយ C) នៃខ្សែសង្វាក់ H នៅជិតស្ថានីយ C និង 1/2 (មានសំណល់អាស៊ីតអាមីណូប្រហែល 105) នៅជិតស្ថានីយ C នៃខ្សែសង្វាក់ L ។តំបន់មុខងារនីមួយៗនៃខ្សែសង្វាក់ H មានសំណល់អាស៊ីតអាមីណូប្រហែល 110 ហើយមានចិញ្ចៀន peptide ដែលផ្សំឡើងដោយសំណល់អាស៊ីតអាមីណូ 50-60 ដែលតភ្ជាប់ដោយចំណង disulfide ។សមាសធាតុអាស៊ីតអាមីណូ និងការរៀបចំនៃតំបន់នេះគឺថេរក្នុងខ្សែសង្វាក់ Ig isotype L សត្វដូចគ្នា និងខ្សែសង្វាក់ H ប្រភេទដូចគ្នា។ដូចគ្នាដែរ វាអាចភ្ជាប់ជាពិសេសទៅនឹងអង់ទីហ្សែនដែលត្រូវគ្នា ប៉ុន្តែរចនាសម្ព័ន្ធនៃតំបន់ C របស់វាគឺដូចគ្នា ពោលគឺវាមានអង់ទីហ្សែនដូចគ្នា។អង់ទីករបន្ទាប់បន្សំ IgG ប្រឆាំងនឹងមនុស្ស (ឬប្រឆាំងនឹងអង្គបដិប្រាណ) អាចត្រូវបានផ្សំជាមួយនឹងការរួមបញ្ចូលគ្នានៃអង្គបដិប្រាណ (IgG) ពីរប្រឆាំងនឹង exotoxins ផ្សេងគ្នាកើតឡើង។នេះគឺជាមូលដ្ឋានដ៏សំខាន់សម្រាប់ការរៀបចំអង្គបដិប្រាណបន្ទាប់បន្សំ និងអនុវត្ត fluorescein អ៊ីសូតូប អង់ស៊ីម និងអង្គបដិប្រាណដែលមានស្លាកផ្សេងទៀត។
ផលិតផលដែលពាក់ព័ន្ធ៖
ឧបករណ៍ Cell Direct RT-qPCR
ពេលវេលាប្រកាស៖ ថ្ងៃទី ៣០ ខែកញ្ញា ឆ្នាំ ២០២១



